|
|||||||||||||
| Home | Research | People | Publications | BBS | Photos | Contact Us | |||||||
|
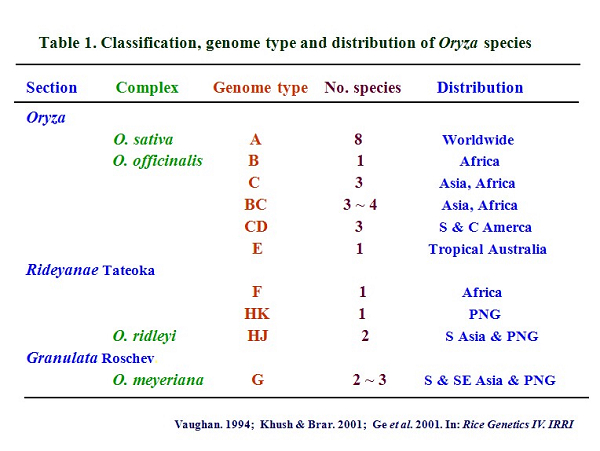
1. Molecular phylogenetic reconstruction, with special emphasis on the rice tribe and genus It is well known that rice is one of the world's most important crops, and is becoming an excellent model for various biological studies, in particular after the completion of the rice genome projects. The rice genus consists of 23 wild species and includes important genetic resources in rice breeding and improvement (Table 1). Taking full advantage of rice genetic and genome resources relies significantly on a clear understanding of the evolutionary relationships of rice and its relatives.
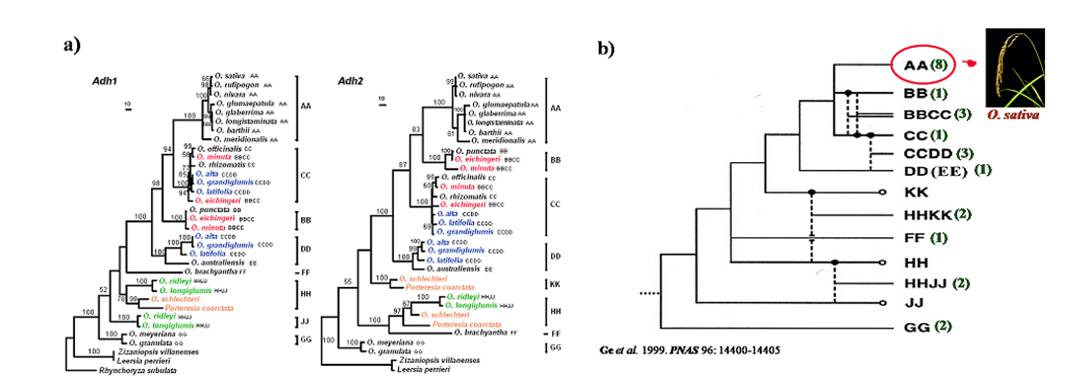
In previous studies, we established a good phylogenetic framework of the rice genus (Figure 1-1): 1) in addition to supporting previous recognition of nine genome types (A, B, BC, C, CD, E, F, G, and HJ) in the rice genus, we recognized a new genome type, HK, for Oryza schlechteri and Porteresia coarctata, suggesting that P. coarctata might be included in Oryza; 2) the E-genome species was most closely related to the D-genome progenitor that gave rise to the CD-genome species; 3) In contrast to a single origin of three CD-genome species, the BC-genome species had different origins because their maternal parents had either a B- or C-genome; 4) The G genome is the most basal lineage in the rice genus and the A-genome group, which contains the two cultivated rice species, was a recently diverged and rapidly radiated lineage within the rice genus (Ge et al. 1999. PNAS).
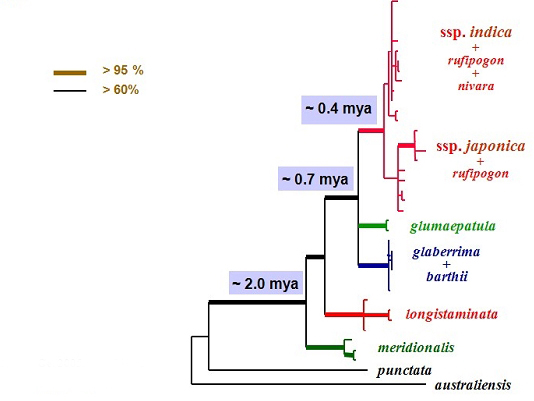
Figure 1-1. Phylogeny of the Oryza species. A) phylogenetic trees based on Adh1 and Adh2 genes. Numbers are bootstrap percentages above 50%. B) Evolutionary relationships of the rice genomes inferred from Adh1, Adh2, and matK genes phylogenies. Broken lines indicate origins of allotetraploids. Solid and open circles indicate maternal parents and unidentified diploid genomes, respectively. Letters stand for the genome types and figures in parentheses indicate the number of species. We also reconstructed the phylogeny of the A-genome species that included the cultivated rice (Figure 1-2) (Zhu & Ge 2005. New Phytol). The resulting phylogeny demonstrated that the Australian endemic O. meridionalis was the earliest divergent lineage and supported previous opinion to treat O. rufipogon and O. nivara as a single species. More importantly, we indicated that two subspecies of O. sativa (ssp. indica and ssp. japonica) formed two separate monophyletic groups, suggestive of their independent origins from two ancestor lineages that separated from each other about 400,000 years ago (Zhu & Ge 2005. New Phytol).
Figure 1-2. Phylogeny of the Oryza species with A-genome type. Shaded figures indicate the divergence times for the branches in million years ago (mya). We then investigated the phylogenetic relationships between the rice genus and its closely related genera (Ge et al. 2002. AJB; Guo & Ge 2005. AJB). Based on chloroplast, mitochondrial and nuclear genes, we sampled all 12 genera in the rice tribe, and revealed that 1) the rice tribe formed two monophyletic groups corresponding to the traditionally recognized subtribes; 2) Leersia is the most closely related genus to Oryza and the two genera separated about 9 mya; 3) previous recognition of three monotypic genera (Porteresia, Hydrochloa, and Prosphytochloa) is not justified; 4) close affinities of the monoecious genera are not supported, suggesting the possibility of multiple origins of unisexual florets? (Guo & Ge 2005. AJB). Accordingly, we revised the taxonomic treatment of the tribe and returned to Oryza the genus Porteresia that is an important germplasm for salt tolerance (Lu & Ge 2005. Nord J Bot ).
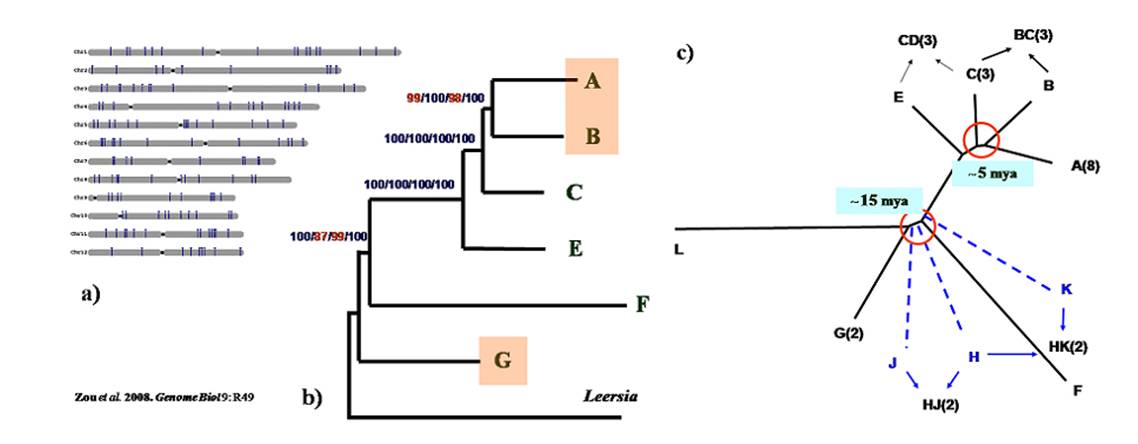
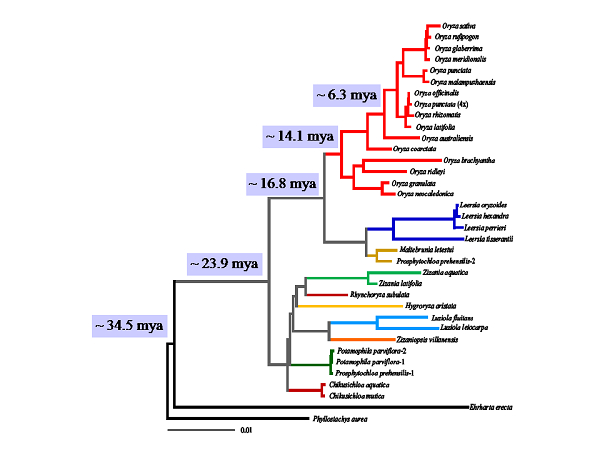
Figure 1-3. A phylogenomic study revealed rapid radiation of the Oryza species. a) Relative physical locations of the 142 sampled genes on rice chromosomes. b) A fully resolved phylogenetic tree of diploid genomes in Oryza based on the concatenated 142 genes. Numbers above branches indicate bootstrap support of ML, MP, and NJ, and posterior probability of BI, respectively. C) A schematic illustration of two episodes of rapid speciation that gave rise to almost the entire diversity of the rice genus. Letters represent the genome types and figures in parentheses indicate the number of species. We further generated a fully resolved phylogeny of Oryzeae including all extant genera in the tribe based on the sequences of 20 chloroplast fragments (Figure 1-4). We revised the systematic position of Prosphytochloa and suggested that the previously unresolved phylogeny of the subtribe Zizaniinae was most likely explained by insufficient data due to rapid radiation. We estimated the crown node of Oryzeae and the deepest split of Oryza at about 24 and 15 MYA, respectively. We developed a biogeographic history of the tribe and substantiated the important role of long-distance dispersal in the origin and diversification of the tribe Oryzeae (Tang et al. 2010. MPE).
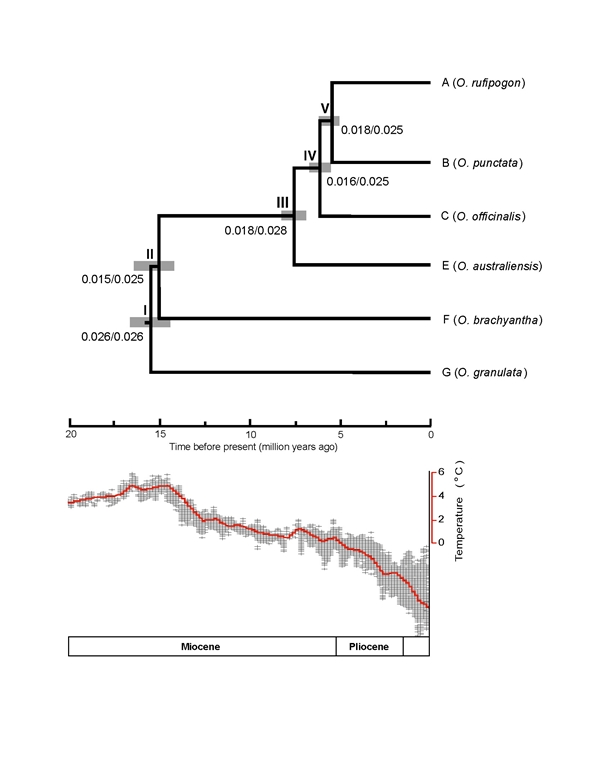
Figure 1-4. Phylogeny and divergence time of the main lineages in the rice tribe, inferred from the concatenated 20 chloroplast fragments. Numbers on branches indicate the divergence time in million years ago (mya). Using phylogenomic approaches, we explored the evolutionary radiations in Ehrhartoideae at different taxonomic levels. We estimated two radiation events happened in the evolutionary history of Oryza, with the first in the middle Miocene (c. 13-15 Ma) shortly after its origin, and the second c. 5-6 Ma. These events gave rise to the whole genome diversity in this genus. We suggest that the climate fluctuations may have contributed to the two rapid diversifications of Oryza species (Figure 1-5) (Zou et al. 2013. New Phytol). In addition, analyses of chloroplast genomes suggest an earlier radiation in the initial diversification of the BEP clade into three subfamilies (Bambusoideae, Ehrhartoideae and Pooideae) (Wu & Ge 2012. MPE).
Figure 1-5. The chronogram of the genus Oryza based on 106 loci. Below the chronogram is the figure showing temperature changes since the Miocene.
|
| © The Ge Laboratory Institute of Botany, the Chinese Academy of Sciences |